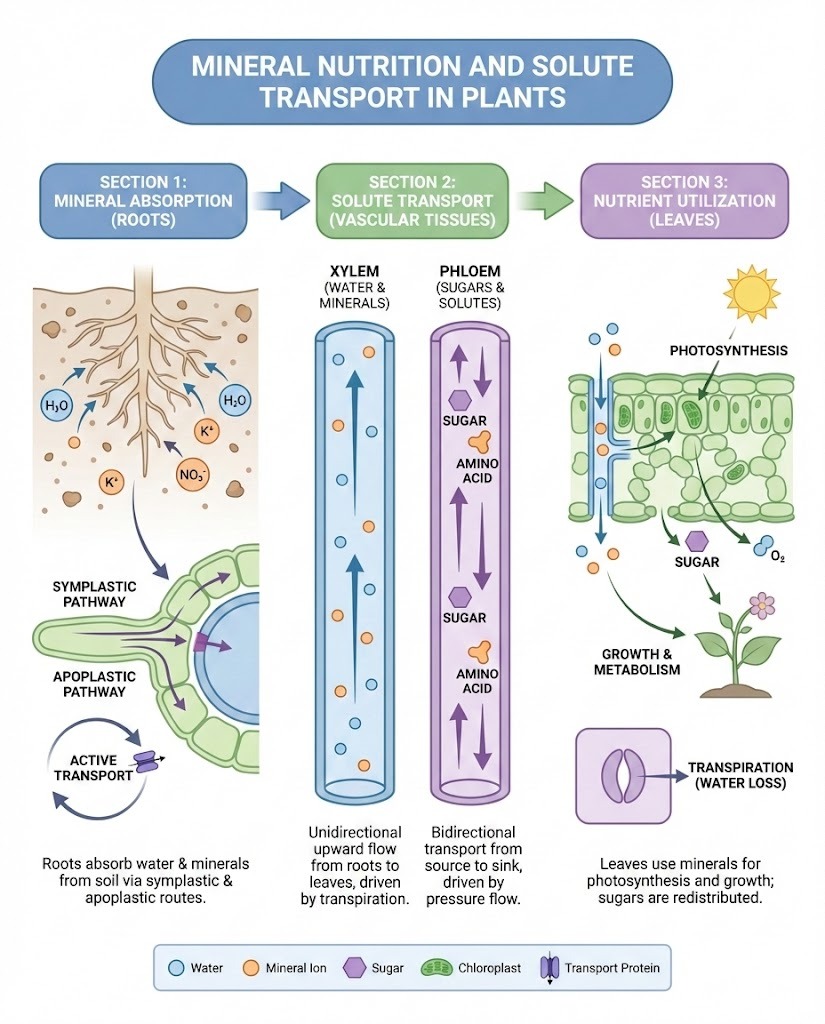
Plant life is sustained by a process of mineral nutrition, whereby essential chemical elements are absorbed from the environment to support growth, development, and reproduction. While plants obtain carbon and oxygen from the atmosphere, and hydrogen from water, the remaining thirteen or more mineral nutrients are primarily absorbed from the soil solution through the plant roots in their ionized forms. These essential elements are divided into macronutrients, which are required in relatively large quantities (such as Nitrogen, Phosphorus, and Potassium), and micronutrients or trace elements, which are needed in much smaller amounts (like Iron, Manganese, and Zinc). Both groups are indispensable, serving roles from providing structural strength to cell walls and forming the core of the chlorophyll molecule (Magnesium) to activating critical enzymes in metabolic processes. The availability of these nutrients in the soil in a soluble, ionic form is therefore a prerequisite for healthy plant growth.
Solute Transport Across the Plasma Membrane
The initial and most fundamental barrier for any nutrient entering a plant cell is the plasma membrane. Being a lipid bilayer, this membrane is, in principle, impermeable to polar molecules and charged ions, meaning all solutes must be transported by specialized proteins embedded within the membrane. This transport is categorized into two major mechanisms: passive transport and active transport. Passive transport, which includes diffusion and facilitated diffusion (mediated by channels or carrier proteins), moves solutes down their concentration or electrochemical gradient and does not require the direct expenditure of metabolic energy (ATP). Conversely, active transport moves ions and molecules against their concentration gradient, which is an energy-consuming process. This energy is typically supplied either directly by the hydrolysis of ATP (primary active transport, e.g., H+-ATPase proton pumps) or indirectly by coupling the movement of the solute to the movement of an ion (like H+) down its electrochemical gradient, a process known as secondary active transport or cotransport. This active, selective uptake is crucial because the concentration of essential ions is often much higher inside the root cells than in the soil solution, requiring an uphill movement.
Radial Transport Pathways in the Root
Once absorbed at the root surface, nutrients are transferred radially towards the central vascular tissue (stele) using three interconnected pathways. The apoplastic pathway involves the movement of water and dissolved solutes through the non-living spaces of the root—primarily the cell walls and intercellular air spaces—without ever passing through a cell membrane. This is a bulk, relatively unregulated flow. The symplastic pathway, or cell-to-cell pathway, begins when a nutrient crosses the plasma membrane of an epidermal cell (often a root hair) and then travels continuously from the cytoplasm of one cell to the next through the cellular bridges called plasmodesmata. The symplast forms a living cellular continuum throughout the root cortex. A third route, sometimes considered a component of the symplast route, is the transcellular pathway, which involves solutes repeatedly crossing the plasma membrane to enter and exit adjacent cells. The symplastic route is highly significant because the initial step of crossing the plasma membrane allows the cell to selectively filter and control which substances are permitted to enter the living tissues of the plant.
The Endodermis: A Critical Transport Barrier
The selective process of nutrient uptake reaches a decisive point at the endodermis, a cylinder of cells that surrounds the vascular tissue of the root. Cells of the endodermis possess a water-impermeable band of suberin called the Casparian strip in their radial and transverse walls. This suberinized structure acts as a critical transport barrier, completely blocking the apoplastic movement of water and solutes at this layer. Consequently, any substance that has been traveling apoplastically must, at this junction, be shunted into the symplastic pathway by passing through the plasma membrane of an endodermal cell. This mechanism ensures that all essential mineral nutrients must pass through at least one living cell membrane before reaching the stele, thereby enabling a final, rigorous selection and filtration step that prevents the uncontrolled flow of toxic or unwanted ions into the plant’s long-distance transport system. Following this filtration, nutrients are often actively accumulated by xylem parenchyma cells to a high concentration before being loaded into the xylem vessels.
Long-Distance Transport in Xylem and Phloem
Plants utilize two specialized vascular tissues for transporting solutes over long distances. The xylem is responsible for the bulk, unidirectional upward flow of water and dissolved mineral ions from the roots to the shoots and leaves. This flow is primarily driven by transpiration—the evaporation of water from the leaf surface—which creates a negative pressure (tension) that pulls the water column upwards, a phenomenon known as the cohesion-tension theory of sap ascent. The phloem, in contrast, serves to translocate organic solutes, predominantly photosynthetically produced sugars (sucrose), from their source (e.g., mature leaves) to sink tissues (e.g., roots, developing fruits, or growing tips). Phloem transport is bidirectional and is driven by a pressure flow mechanism resulting from the osmotic gradient created by the active loading of sugars at the source. Furthermore, the phloem plays a significant role in the redistribution of mobile mineral nutrients, such as Nitrogen and Potassium, from older, senescing tissues to younger, actively growing parts of the plant, maximizing nutrient use efficiency.
The Role of Transpiration in Mineral Supply
A long-standing assumption in plant physiology is that transpiration is necessary for the long-distance transport of mineral nutrients in the xylem. While the increased flow of water during high transpiration undoubtedly accelerates the rate at which dissolved mineral ions move upward—a process called mass flow—experimental evidence suggests this acceleration is not strictly essential for plant viability or growth. Studies that reduced the rate of transpiration significantly (e.g., by supplying nutrients during the night or growing plants under high humidity) demonstrated that mineral uptake and growth were often unaffected. This indicates that other forces, specifically convective water transport mechanisms such as root pressure (generated by the active accumulation of ions into the xylem) and water flow driven by growth, are in themselves sufficient to deliver an adequate supply of minerals for plant function. Thus, while transpiration is the major driver for bulk water flow and rapid nutrient movement, its absence does not preclude the essential delivery of minerals, highlighting the robustness of the plant’s internal transport architecture.
Interconnected Solute Transport and Cellular Homeostasis
The overall system of mineral nutrition and solute transport in plants is characterized by an intricate and non-isolated network of processes. It combines passive and active membrane transport, selective radial movement through the root, and highly efficient long-distance vascular transport. The integrity of this entire system is essential for maintaining the plant’s cellular integrity, turgor, and chemical homeostasis. The selective barriers, the specialized transport proteins, and the dual vascular conduits work in concert, ensuring that the plant not only acquires the necessary chemical building blocks from the soil against unfavorable concentration gradients but also efficiently distributes and redistributes them throughout its structure to support sustained growth and development.